Thursday, July 05, 2007
Bright Eyes

Deer apparently have reflectors for eyes.

(Same pic with brightness boosted. This is my front yard.)
The prevailing theory of visual cortex assumes that cortical mini-columns, which are anatomically distinct, vertically-aligned clusters of ~100 neurons, effectively act as unit, with all of the cells in the column working together to detect and represent some preferred (by that column) feature. This is a fairly empirical result, based on presenting various stimuli to live animals and observing that the cells within a mini-column tend to fire together.
A study[1] I found a few years ago, however, showed that if you simply pay attention to enough aspects of the input, the individual cells within the mini-column appeared to represent quite distinct features from eachother--i.e., the column doesn't act as a unit, it only looks that way because we're presenting stimuli that are too vague to show up the differences. In fact the cells appear to encode roughly orthogonal features to each other, which is a good thing from an information-theoretic perspective. (Though it's a bad thing from a computational perspective because it means a hundred times as many distinct units to simulate.)
A more recent paper[2] with similar results noted:
These differences in [feature] space may not be apparent when probed with simple stimuli that vary in only one or two stimulus parameters. Neighboring neurons may appear to exhibit very little response heterogeneity in that case, simply because the full range of stimulus dimensions is not being implemented. Only when probed with rich spatio-temporal visual stimuli, as is the case with natural movies, will the true heterogeneity of the local population be revealed.
In other words, overly simplified images in, overly simplified neural activations out. Present real-world, moving images, and presto, the cells in the mini-column start firing sparsely and with high contrast:
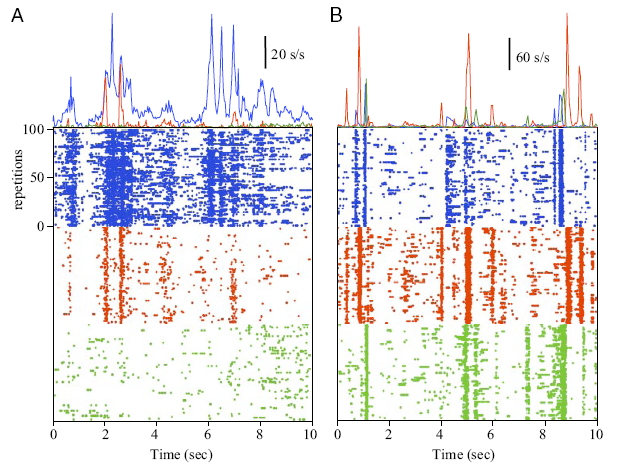
"Peristimulus time histograms (PSTHs) and raster plots of the spike times recorded from a single tetrode in 2 different experiments (A and B) to 100 repetitions of a 30-s movie (10 s of data are shown). In both sets of recordings, the activity of 3 single units are displayed. As in Fig. 1, the activities of cells 1-3 are shown in blue, red, and green, respectively. Responses to specific frames of the movie are apparent from the alignment of spikes across repetitions and the well-defined changes in average firing rate in the PSTHs. These data illustrate that cells recorded on the same tetrode often exhibit clear differences in firing rate, response reliability, and time course."
Not to miss in the above plot is that each of those color bands represents a single neuron's response to multiple repetitions of the same few seconds of a movie (each repetition is another row of pixels of the same color, all stacked vertically to make the thick band). Look at the repeatability (reliability) of those responses! And concurrently, note that the various (near-by) cells are responding very differently from each other. It looks like a pretty concrete, usable encoding--strikingly non-nebulous. (Unfortunately, they didn't have enough data to infer the visual features preferred by each of those cells. That would have been really cool.)
Another interesting result from the same paper:
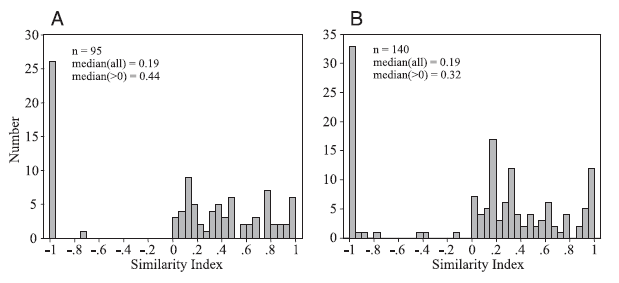
"Histograms of the similarity index for cell pairs recorded on the same (A) and different (B) tetrodes."
Ignore the A/B comparison--it suffices to look at either on its own for this point: This is a histogram of correlations between pairs of cells. Of note is that many pairs have fairly random positive correlations (tend to fire together more often than chance, but not strikingly so), but on the negative side, there's a valley and then a huge spike at the negative extreme (which by their metric means the cells essentially never fire together). I don't recall them explicitly commenting on this, but to me that graph has interesting implications about lateral inhibition--it seems to imply that if two cells are connected by an inhibitory interneuron, they're pretty much hard-wired never to fire (burst, really) at the same time. I.e., no learning, no grey zone, just declare these two cells mutually exclusive. It's possible (likely) that there are other styles of (learned) lateral inhibition going on as well, but this chart sure implies a hard form is strongly in play. [There's nothing really new in that, but I find this empirical histogram nicely illustrative.]
[1] Functional Micro-Organization of Primary Visual Cortex: Receptive Field Analysis of Nearby Neurons; Gregory C. DeAngelis, Geoffrey M. Ghose, Izumi Ohzawa, and Ralph D. Freeman
[2] Heterogeneity
in the Responses of Adjacent Neurons to Natural Stimuli in Cat Striate Cortex
Shih-Cheng Yen, Jonathan Baker and Charles M. Gray; J Neurophysiol 97:1326-1341, 2007